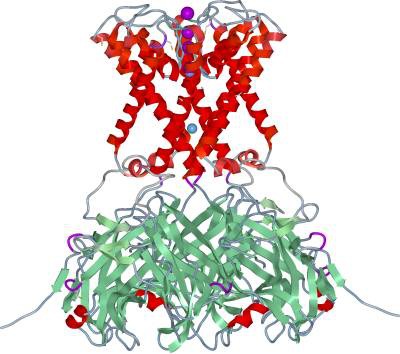
All channels exist in two major conformations: the closed and open states.
Tight regulation of the transition process from the closed to the open state,
known as gating, is key to ensuring that channels do not
kill cells by dissipating their ionic gradients.
However, equally as important is the fact that channels
have to respond quickly and efficiently to a signal.
There are many different signals that channels respond to, including
the change in voltage across the membrane and the binding
of an assortment of ligands such as ATP, cAMP, ions and/or lipids.
In order for the channel to respond to the appropriate ligand,
they have specific regulatory domains attached to the ion conduction pathway.
The large number of regulatory domains ensures that a family of ion channels
such as the K+ channels can be involved in a wide range of physiological processes
from the release of hormones to the generation and propagation of electrical signals.
The two crystal structures of the K+ channel KirBac3.1 from
Magnetospirillum magnetotacticum reveal important structural
details of the gating process as the channel tranforms from the closed toward the open state.
KirBac3.1 is a member of the inwardly rectifying K+ (Kir) channel family,
a ligand-gated ion channel.
The structures are designated intermediate states 1 and 2 (IS1 and IS2).
In comparison with the closed
KirBac1.1 structure, a significant difference was observed in the arrangement
of ions within the selectivity filter.
This region, located on the extracellular side of the membrane,
selects for K+ ions over the other monovalent cations.
There are potentially four ion binding sites within the selectivity filter,
designated S1 to S4.
In the IS1 structure three K+ ions are found in sites S1, S2 and S4.
The ion arrangement in the selectivity filter shifts more toward the open state
in the IS2 structure with all four sites occupied.
However, there is a significant aniostropic displacement of the ions
around their respective binding sites in IS2.
Kir channels obtain their name from the fact that they
preferentially allow the movement of ions in the inward direction.
This is due to the outward movement of K+ ions being blocked
by other cations that do not permeate the entire ion conduction pathway.
It is known that both Mg2+ and polyamines are responsible for blocking K+ ion movement.
A new Mg2+ binding site has been identified in both intermediate states.
The location is in the selectivity filter on the side facing the cytoplasm,
an ideal position for blocking K+ ion movement.
The central cavity in all K+ channels helps to stabilize an ion
in the centre of the membrane by surrounding the ion
with a polarisable medium - water.
Previously, it was believed that the water was unordered.
However, the KirBac3.1 structures reveal a monolayer of water in the cavity.
In the centre where the ion conduction pathway exists the waters are in a four-fold arrangement.
This arrangement mimics the selectivity which has a four-fold arrangement of carbonyl oxygen atoms.
Hence it is believed that the waters in the cavity form an extension
of the selectivity filter potentially directing ion flow.
In the closed state of KirBac1.1, four hydrophobic Phe residues,
located on the inner helices at the membrane/cytoplasm interface,
come together to prevent ion movement indirectly by blocking water movement.
As the channel goes into the open state these residues have to move,
freeing the ion conduction pathway.
In the IS2 structure, its inner helices have partially opened up.
This time the space in between the helices form a binding site for the second type of blocker,
the polyamine spermine.
This flexible molecule is held in place by hydrogen bonding
to Tyr residues and a cation/pi interaction.
Overall the two KirBac3.1 crystal forms shed light on the
intricate process of an ion channel as it progresses
from the closed state toward the open state.
The structures reveal changes in ion binding within the selectivity filter
and movements of the inner helices.
Mg2+ and spermine binding sites have been identified
and the waters in the central cavity appear to be playing a
more significant role in the process of ion conduction.